3 Главный комплекс гистосовместимости
Главный комплекс гистосовместимости - комплекс тесно связанных генетических локусов, расположенных на коротком плече 6-ой хромосомы (6р), а также их белковых продуктов, отвечающих за развитие иммунного ответа и синтез трансплантационных антигенов. Этот комплекс содержит больше 200 генов[1]. Эти гены отличаются высокой полиморфностью, кодоминантностью[2]. В базе данных IPD-IMGT/HLA на 27 февраля 2023 года содержится информация о 35820 человеческих аллелях HLA[3]. Популяции по генам МНС имеют большой процент гетерозиготных особей[4]. Сцепленные группы генов МНС называются гаплотипами. Функции генов МНС заключается в следующем (жирным выделены функции, обозначающие цель существования такой системы):
Обеспечение процессинга и презентации антигенных пептидов - индукторов и мишеней иммунного ответа;
Обеспечение взаимодействия клеток
Распознавание собственных, измененных собственных и чужеродных клеток; запуск и реализация иммуного ответа против носителей генетической информации
Поддержание иммунологической толерантности (в том числе во время беременности);
Участие в селекции Т-лимфоцитов;
Создание генетического разнообразия и обеспечение выживаемости вида.
Все гены локуса МНС поделены на три класса: первый, второй и третий. На Figure 3.1 представлена генетическая карта генов МНС[5].
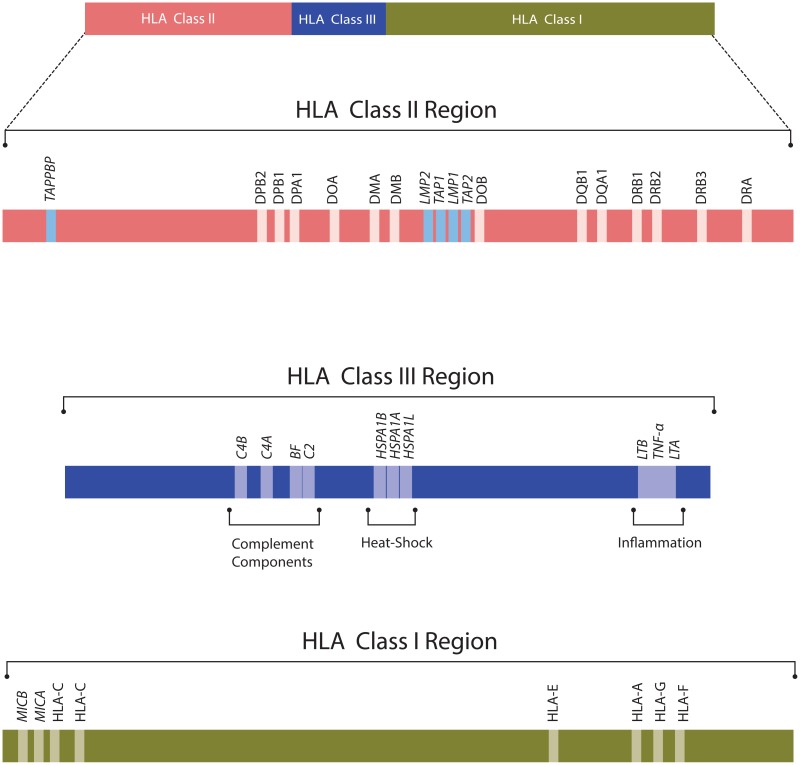
К классическим генам относят три пары генов, кодирующих \(\alpha\)- и \(\beta\)- цепи МНС второго класса: HLA-DR, HLA-DP и HLA-DQ - и три гена кодирующие \(\alpha\)-цепь МНС первого класса: HLA-A, HLA-B, HLA-C. Зрелые продукты этих генов располагаются на цитоплазматической мембране и непосредственно участвуют в презентации антигенов. У многих людей кластер HLA-DR содержит дополнительный ген \(\beta\)-цепи, который может соединяться с DR\(\alpha\)-цепью[6]. Помимо этого существует большое количество генов “неклассических” МНС, которые по структуре напоминают гены МНС, но их продукты не презентируют пептиды обычным \(\alpha\beta\)Т-лимфоцитам, но выполняют ряд других функций в клетке[6]. Например, MIC A, MIC B ответственны за обеспечение контакта с NK-клетками, \(\gamma\delta\)Т-лимфоцитами, HLA-DM и HLA-DO участвуют в процессинге антигенов для HLA второго класса[7; 8], HLA-G подавляют подавляют способность цитотоксических лимфоцитов секретировать IFN-\(\gamma\) и стимулируют секрецию TGF-\(\beta\), что играет важную роль в обеспечении толерантности матери к антигенам плода[9]. В локусе генов МНС содержатся гены, чьи продукты прнимают участие в процессенге антигенов для MHC первого класса (TAPBP, TAP1,TAP2, LMP1,LMP2). Выделяют особую группу генов МНС третьего класса, продукты которых не участвуют в презентации антигенов, но играют важную роль в обеспечении иммунитета[10]. Это компоненты комплемента (С4А, С4В, С2), белки теплового шока (HSPA1A, HSPA1B), белки семейства фактора некроза опухоли (LTB, LTA, TNF-\(\alpha\)).
Рассмотрим строение главных комплексов гистосовместимости первого и второго класса. МНС первого класса - гетеродимер, состоящий из большой \(\alpha\)-цепи, которой он заякорен в цитоплазматической мембране, и \(\beta_2\)-микроглобулина. \(\alpha\)-цепь имеет 3 домена. \(\alpha_1\)- и \(\alpha_2\)-домены формируют особый сайт - пептидсвязывающую бороздку, которую иногда в литературе называют в честь учёной, впервые описавшей структуру МНС I и МНС II, по фамилии Бьоркман[11]. Ген \(\beta_2\)-микроглобулина не относится к генам МНС и расположен на 15 хромосоме. \(\beta_2\)-цепь связана с \(\alpha\)-цепью нековалентными взаимодействиями. Этот комплекс нестабилен, пока в пептидсвязывающей бороздке нет пептида. В ЭР он стабилизируется шапероном кальнексином. Комплексы, которые презентируют слабоаффинные эпитопы, быстро диссоциируют на ЦПМ, и цепи возвращаются обратно в ЭПР благодаря реакциям белкового разрушения, связаным с ЭР (endoplasmatic reticulum-associated protein degradation, ERAD)[12]. Геометрия бороздки Бьоркман у МНС первого класса “закрытая”, поэтому в неё помещаются пептиды ограниченного размера, длиной 8-10 аминокислот[13]. МНС второго класса представляет из себя гетеродимер. Обе цепи, \(\alpha\) и \(\beta\), имеют по два домена и связаны между собой нековалентными связями. Пептидсвязывающую бороздку формируют \(\alpha_1\) и \(\beta_1\) домены[14], и она, в отличии от МНС первого класса, “открытая”, поэтому в неё помещаются более длинные пептиды. Обычно длина пептидов, презентируемых на МНС второго класса, колеблется от 12 до 25 аминокислот[15]. Как и МНС первого класса, МНС второго класса нестабилен без эпитопа.
Структуру пептидсвязывающей бороздки у МНС первого класса можно разделить на 6 карманов, причем крайние карманы А и F отличаются консервативностью[16]. Поэтому для связывания с МНС первого класса важную роль играют N-концевые и C-концевые остатки, которые связываются с крайними карманами. От характера величины силы нековалентных взаимодействий этих карманов с некоторыми остатками пептида, которые называют якорными, зависит спектр связываемых эпитопов этим МНС. В зависимости от аллеля МНС может быть допольнительные первичные якорные остатки. Между ними могут находиться аминокислотные остатки с сильно варьирующими по физико-химическим свойствами. У МНС второго класса якорные остатки могут быть расположены на различном удалении друг от друга, кроме этого пептид может выходить за пределы пептидсвязывающей бороздки, следовательно, в его связывании также участвуют и другие структуры белка[6].
МНС двух классов презентируют свой пептид разным клеткам: МНС первого класса презентирует пептиды, распознаваемые CD8+ лимфоцитами, а МНС второго класса - CD4+ лимфоцитами[17]. МНС первого класса экспессирован практически на всех ядросодержащих клетках организма[18]. Единственное исключение - клетки ворсинчатого трофобласта. Показано, что экспрессия классических аллелей МНС I на них приводит к иммуноопосредованному аборту[19]. МНС второго класса экспрессируются конституциально, клетками иммунной системы, в особенности антигенпрезентирующими клетками: дендритными клетками, макрофагами, В-лимфоцитами, эпителиальными клетками тимуса, но их экспрессия может быть индуцирована IFN-\(\gamma\) на остальных клетках[20].
От локализации патогена зависит, на каком МНС будут презентироваться его антигены. Например, разрушенные белки цитозольных патогенов - вирусов, некоторых бактерий - транспортируются в ЭПР и там встраиваются в вновь синтезированные МНС первого класса и оттуда готовые комплексы доставляются на цитоплазматическую мембрану[21]. Такой каскад реакций называют непосредственным презентированием. При непосредственном презентировании белки разрушаются специальным белковым комплексом - протеасомой. Помимо непосредственного презентирования существует каскад реакции, когда экзогенные пептиды презентируются на МНС I. Существуют патогены, способные ненапрямую поражать АПК. Существует специальный тип дендритных клеток, который презентирует пептиды, полученные фаголизосомальной системой, на МНС первого, а не второго класса, как это протекает в других АПК[22]. Такой каскад называется перекрестной презентацией, а активацию Т-клеток, с помощью этого каскада - кросс-праймингом. Рассмотрение перекрестного презентирования и презенитрования на МНС II выходит за рамки данные работы, поэтому далее речь пойдет о процессах протекающих при непосредственной презентации антигена.